To support my work: https://www.patreon.com/statedclearly
To support ClockWork Biology: https://www.patreon.com/clockworkshow
Special thanks to @Clockworkbio and for @Spronk3d for their extremely useful protein animations.
How do proteins really work and evolve? This video is where you'll find out!
LEARN MORE:
To see David Goodsell's interactive protein tour, go here: https://cdn.rcsb.org/pdb101/molecular-machinery/
To see David's amazing biochemistry art, go here: https://pdb101.rcsb.org/sci-art/goodsell-gallery
FURTHER READING:
Brownian Motion at the level of proteins: https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1000694
How Lysozymes work as "Brownian motors": https://pmc.ncbi.nlm.nih.gov/articles/PMC4916960/
How Lac Repressor works: https://pmc.ncbi.nlm.nih.gov/articles/PMC220206/
For protein surface charges I used PyMol. Surface charges were estimated and colored using APBS Electrostatics by:
Dolinsky TJ, Nielsen JE, McCammon JA, Baker NA.
PDB2PQR: an automated pipeline for the setup, execution, and analysis of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Research 32 W665-W667 (2004).
Оглавление (7 сегментов)
Segment 1 (00:00 - 05:00)
If you clicked on this video, you already know that proteins are far more than nutrients found in beans and meat. They are the self-assembling biochemical machines that drive the activities of all living cells. At the basement of life, nature tinkers around with proteins, evolution's squishy, magnetic Legos. Magnetic Legos that evolution has been tinkering with for almost 4 billion years, maybe even longer. Proteins come in many shapes and sizes. Most of the particles you're seeing here in this illustration of a cross-section of E. coli, most of the particles here are proteins. They are packed inside the cell like it's a mosh pit. They bounce around smashing off each other at incredibly high speeds. Even though this looks like chaotic noise, most proteins are functional. Collagen proteins serve a fairly boring but very important structural function inside your skin, muscles, and bones. They form fibers. Other proteins function as molecular machines. They have moving parts. They actively do things either inside or outside the cell. Lysozymes, for example, are little proteins secreted in your tears and in your snot. They are attracted to bacteria like a magnet and they rip them apart on contact. Some proteins act as chemical transporters. Here we are looking at a hemoglobin protein complex. Hemoglobin is the stuff crammed inside your blood cells helping spread oxygen through your body. Each little red guy here is hemoglobin. And here we see an example of what are sometimes casually referred to as computing proteins. They're called that because by binding to DNA under certain specific conditions and releasing DNA under different specific conditions, these proteins function like logic gates. They {quote} compute when a cell should or should not use a specific gene. Last but not least, the bacterial flagellum is a protein complex. It's a bunch of different proteins stuck together and together as a whole, they function as a motor. Okay, but how? Protein is a chemical, right? How does a chemical Well, first of all, how does it exist in so many crazy different forms? Some of these proteins are hundreds of times larger than others. Why do we call them all proteins? What's the common denominator here? How do they perform such dramatically different functions from digestion to scaffolding to computing to swimming? How? How does one thing do so much different stuff? And what about the way they move? When you look at animations of proteins self-assembling, animations like this beautiful one here from Peter over on the YouTube channel Clockwork Bio, or when you see true-to-life simulations showing how proteins move inside the cell, it almost seems like magic. They just fly around the cell like Superman. How does that work? I mean, Superman can fly because he's imaginary. Proteins are supposedly real. What's going on here? After today's lesson, you will be able to intuitively understand physically how these molecular machines function. You will understand how they automatically fold as they are built by the cell. You will understand the main energy source that powers their movements. And last but not least, you will understand how protein complexes A protein complex is a group of proteins attached together. You will understand how protein complexes self-assemble. Proteins, as spectacular as wonderful as magical as they might seem, they can be studied. They can be understood. I am John Perry from Stated Clearly. You are watching How did bacterial flagella evolve? Episode 6, Protein, evolution's fantastic, squishy, magnetic Legos. The first thing you need to know to understand how proteins work is that proteins are strings. This is so important that it's the reason I built this little homemade model here. It's the reason I went through all the trouble. Proteins, believe it or not, they are strings. Even though we always see them depicted as globs, as these three-dimensional shapes, they are actually strings that are bent and folded in on themselves. What are proteins strings of? Proteins are strings of amino acids. Here is what amino acids look like in atomic form if you actually show each atom in the amino acid. There are 20 standard types of amino acids commonly found in proteins. Each type is fascinating, but we only need to know a few things about amino acids in general to understand how proteins work. First, all amino acids share a nearly identical backbone. Here's what the backbone looks like. You can see this amino acid has this little backbone on the top. And the same is true for all the other amino acids. All of them except for proline, but
Segment 2 (05:00 - 10:00)
we're not going to bother talking about proline today. The second really important thing to understand about amino acids is that even though they all share the same backbone, each of the 20 different types of amino acids has a different type of side chain. When we say there are 20 different types of standard amino acids, what we actually mean is there are 20 different standard types of side chains. The amino acid glycine is the simplest. It just has a little hydrogen atom for its side chain. It's basically just a backbone. Tryptophan, in contrast, has a spectacular pair of carbon-based rings as its side chain. It's a huge amino acid. The backbone is dwarfed by the side chain, so much so that you could almost be forgiven for not noticing it even has a backbone at all. Amino acids vary in shape and size because of their different side chains, but those side chains can have other properties as well, aside from just shape and size. Most importantly, some side chains are positively charged or partly so. They're polar. Some are negatively charged or partly so, and others are hydrophobic. What that means is that they are oily, you could say. They are repelled by water like oil is repelled by water. A cell builds a protein, and remember, a protein is a long chain of amino acids. A cell builds a protein by grabbing amino acids and fusing their backbones end to form that chain. Because some amino acids have charges and some are hydrophobic, what this means is that as the protein is being built in the salty water inside of a cell, the protein chain will automatically bend and fold and stick to itself according to the amino acid sequence. As a general rule, as the protein chain is growing, water will push the oily parts, the hydrophobic amino acids, it'll push those towards the center of the glob. That's why it forms these globular structures, part of why it forms globular structures. And also, the positively charged amino acids will be attracted to the negatively charged amino acids because opposites attract. I say that proteins act like magnets, but they're not really magnetic. We're talking about electrostatic charge. Positive charges and negative charges interact with each other at a distance like the north end and the south end of a magnet. Also, very similar to magnets, if you have two positively charged amino acids, they will repel each other. The end result is that a specific sequence of amino acids will always fold into the same 3D shape. There are exceptions to this rule, but almost always. If you were to grab the ends of a folded protein and pull them apart, then let go again in water, it would snap together to form the exact same shape it had before, the exact same three-dimensional shape that it had before. The 3D shape of a folded protein is determined by the sequence of amino acids because of how those amino acids interact with water and how they interact with each other. There are certain proteins that they actually have several different ways that they can fold. And then there are some proteins that don't have a permanent folded structure at all. They're very loose. They kind of wiggle around in the cell like a noodle. But most protein sequences, they only have one way in which they will fold and they fold that way every time the protein is made. If you are new to all of this, what I just told you probably sounds wild, but hopefully it does make sense. Proteins are chains. They automatically fold when wet. If you pull one apart, it will snap back together. Many of the bonds that form between amino acids, they're pretty weak bonds. They're what we call hydrogen bonds. And the reason that I modeled them with these magnets is because just like magnets, if you pull hard on them, they will come apart. That said, if you have a bunch of hydrogen bonds together, a protein will actually be very, very stable cuz lots of those small, weak bonds, they add up to make a strong, stable structure. In most proteins, some regions are highly stable. There's lots of hydrogen bonds holding them together, hydrogen bonds and other bonds. And then other regions are flexible. You can have things that flop around. even spin. And those can have reactive ends on them. When I say that proteins are evolution's squishy, magnetic Legos, this is what I mean by squishy. There are parts of them that move a lot. Parts are stable. Parts can be very stable. In fact, the cross-linked proteins in your fingernails, the keratin protein, they're very, very hard, very stable. But most proteins have squishy parts. They have parts that bend and flex and even spin. Something else worth stating clearly, and this is probably obvious, but the longer an amino acid chain is, the larger the protein will be once it folds up into its three-dimensional shape. Some proteins are just a few dozen amino acids long. Some are thousands of amino acids long. That's why proteins vary so dramatically in size and in shape. So, to sum things up here for now, the size of a protein is determined by the length of the amino
Segment 3 (10:00 - 15:00)
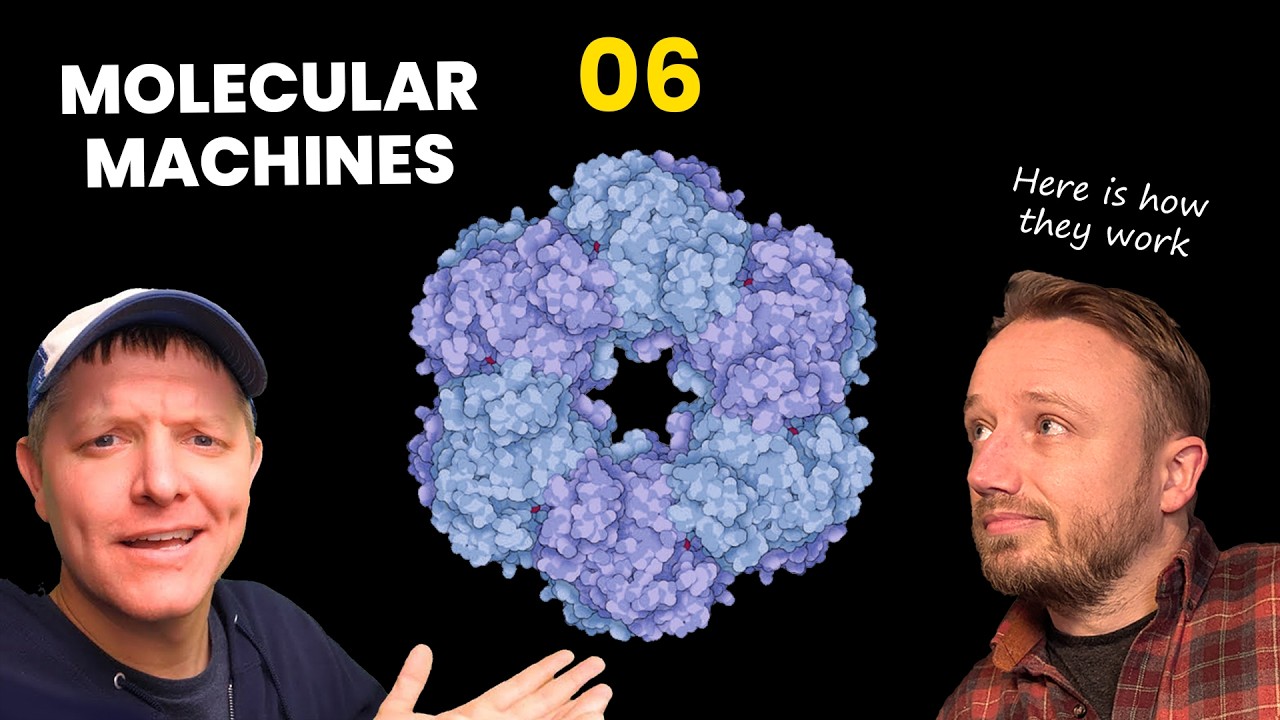
acid chain that makes up that protein. The 3D shape of that protein is determined by the amino acid sequence in that protein chain. All of this is very important to understand, but more importantly still is the fact that the surface of each protein type, once it is folded, has a signature electrostatic charge distribution. That sounds way more technical than it actually is. All I'm saying is that the surface of a protein has parts that are negatively charged, shown here in red, parts that are neutral, shown here in white, and parts that are positively charged, shown here in blue. These surface charges are why I say that proteins are evolution's squishy magnetic Legos. I mean, they're not really magnetic. It is electrostatic attraction and repulsion, but it works similarly to magnetism. Parts of molecules can pull or repel each other at a distance. When you put this all together, it suddenly makes sense why one type of chemical, this thing we call protein, can do so many different things. A protein's size and shape are determined by the number and sequence of amino acids in its chain. Parts of a protein can be highly stable, other parts can bend or spin. Its surface is covered with charges, negative and positive, allowing it to stick to or repel other molecules around it with varying degrees of force. This is actually pretty intuitive, right? The magic behind proteins It can be studied. It can be understood. But we're missing something here, something super important. When we look at simulations of proteins in action, they move. They move a lot, actually. But how? How do they buzz around the cell like Superman? What is the source of their power? At this point you might be thinking, "Oh no, John's going to give us a lesson on ATP, the power currency inside the cell. " Don't worry, we're not going to talk about ATP right now. ATP is a very important molecule, and it is how a lot of proteins power their actions, but it's actually not how most proteins power their actions. Most proteins are simply powered by normal, plain old Brownian motion. Brownian motion, as we all learned back in seventh grade science class, is the random, erratic movement of microscopic particles suspended in liquid or gas. The movement is powered by just the collision of molecules and atoms. The constant, perpetual collision that's happening in any liquid or in any gas. Here I am showing you mud, magnified as high as my little microscope can go. I filmed this myself down in the basement. Because this is mud, when I zoom out, you can see that there are some microbes in here. But the bulk of the background movement, the twinkling you're seeing here, when we're zoomed in, we see that there are minerals fluttering about. That is Brownian motion. It looks like it's alive, but it is not alive. These minerals are being smacked around by water molecules that are bouncing off each other at extremely high speeds due to temperature, a little bit lower than room temperature. Why does this happen? Why does matter constantly jiggle? There's a really cool video by the Slow Mo Guys, Crane Drop in Slow Mo, where they drop 10,000 bouncy balls from a crane. I'm just showing you stills of it here, but there's a moment near the bottom of the screen where the bouncy balls start behaving a lot like molecules in a gas or in a liquid. They're bouncing off of each other. They're colliding in midair. They transfer their momentum one to another. They spin, they rotate. They're just chaotically bouncing off of each other. There's space between them. They're hitting each other and bouncing off. This is what's happening in water. Water molecules are hitting each other. I mean, water molecules, they have a little bit of a charge. The oxygen atom has a slight negative charge. The hydrogen atoms have slight positive charges. And that means that when they pass by each other, they kind of want to stick a little bit when they're bouncing into each other. But in a glass of water like this, these water molecules are constantly bombarding each other. The intensity with which they're hitting each other and bouncing around off each other is directly correlated with temperature. In fact, when we measure temperature, what we're actually measuring is how intense the molecular movement is. If we zoom into a bacterial cell really, really far, this is too zoomed in to film with current microscope technology. So, we're looking at a simulation here instead. Most of the particles you see here are proteins. If you saw last Q& A, you've seen this simulation already. Not only are we super zoomed in, Brownian motion at this scale is incredibly, incredibly fast. In order for us to understand what's happening when we watch this, researchers had to slow it down 3. 3 million times. This entire clip is 6 seconds long, but represents just 1. 8 millionths of a second in real time. I'm playing it on loop for you. The movements inside of a cell, the biochemical are insanely fast. I remember the first
Segment 4 (15:00 - 20:00)
time that I learned about this, when I first learned how fast chemical reactions are, Brownian motion is at the scale of a single protein. I mean, at the scale I can see under my microscope, it's not that fast. But notice, the smaller a particle is, the faster it moves. When I first learned how fast all this was, it blew my mind. It seems impossible. But then I realized, "Wait a minute. My brain works on cellular chemistry. So, if cellular chemistry were slower, my speed of thought would be equally slower, and I would be equally blown away by how fast chemical reactions are. " So, biochemistry is inevitably mind-blowingly fast. It's also worth noting here that gravity does affect proteins. They are influenced by gravity. But because Brownian motion is so intense at just normal biological temperatures, the normal temperatures that a cell runs at, Brownian motion is so intense that gravity essentially doesn't matter inside the cell. Proteins basically live in a zero-gravity mosh pit. One of the things Brownian motion lets proteins do is it lets them, quote, search the cell for partners, and then self-assemble into larger structures once they, quote, find those partners. Check this out. I've made another model for you here. Each of these little black globs represents a protein after it's already folded up. The magnets here represent the charges on the surface of the protein. And this basket with a bunch of just random junk in there, this represents the chaotic clutter inside of a cell. I'm going to shake this thing to represent Brownian motion, and what we're going to watch is we are going to watch these protein complexes self-assemble without any need for divine intervention or any need for an engineer. Just normal Brownian motion and the normal properties of protein allow this to happen. The chemistry itself is competent. It is competent without comprehension. You can see here that there's a little bit of assembly going on. I'm going to speed things up here. Remember, real Brownian motion inside the cell at this scale is millions of times faster than what I'm doing. Even when this is sped up, I'm still millions of times slower with my model here compared to how Brownian motion works in real life. — [snorts] — And look at that, one of them has completely self-assembled. The other one is not finished, as you can see, but if I shake this up just a little bit more, — there we have it. Self-assembly is a thing. Now, the entire process here took 5 minutes. In real life, at the speeds of real Brownian motion, that would have been less than 1 second. But of course, a real cell is a lot bigger compared to its own proteins than my little bin is here. In real life, depending on the concentration of monomers, individual parts, it would probably take these several seconds to assemble if those parts were drifting randomly inside the cell. Self-assembly is amazing, but it's not the only thing that Brownian motion lets proteins do. When you look inside the cell, you see a zoo of proteins. Each one almost seems like its own complex organism, and they're all living in an ecosystem, fighting for scraps, and acting out a struggle for existence of their own. In some ways, that's actually a fairly accurate way to think about them. Proteins have been evolving and interacting with each other billions of years before the first animals ever emerged. Before we had leopards and gazelles, we had lysozymes and hemoglobins. Lysozymes, if you remember from the start of this video, are molecular machines found in human tears that seek out and destroy bacteria. Lysozymes target a tough, sugary fiber that bacteria have, shown here in this illustration, which acts sort of like rebar or internal armor for the cell. It's called peptidoglycan. In some species of bacteria, this sugar is guarded. It's protected by a thick mess of lipids, sugars, and proteins on top of it, as shown here. But that's okay. Mammal tears also have molecules just for dissolving those outer layers, allowing lysozyme proteins to enter and do their deed. The lysozyme seeks out its target through the normal process of Brownian motion. It's just bumping around inside the tears in your eyes until it happens to bump into a bacterium. Let's pretend here that this hand is the lysozyme, and this hand, this arm, is the sugar. If Brownian motion lets these two connect, the surface of the lysozyme is going to attach to the sugar because of its charge. Its charge is the right charge to attach to the sugar. And then, once it's attached, Brownian motion, instead of moving it around is going to cause it to clamp down tight on the sugar. When it clamps sugar, a chemical reaction will occur in
Segment 5 (20:00 - 25:00)
the core of the lysozyme. That chemical reaction will cause the sugar to just break apart. So, just imagine I mean, my special effects budget is not high enough for this, but just use your imagination here. Imagine my hand breaking off, just shooting off the end here during this chemical reaction. The power of that chemical reaction causes the lysozyme to slide down the chain, down that sugary chain. Brownian motion causes it to open again, which resets the chemical reaction. Brownian motion again will cause it to clamp down a second time, and then the chemical reaction will happen again. It will further break the sugary chain, and the power from that reaction will cause the lysozyme to go to the next joint in the sugar. And it will keep on doing that until the sugar is digested sufficiently that the cell just bursts open. That's how the lysozyme works. People call it a Brownian motor. It's not a true Brownian motor. Brownian motion does most of the work, but it's the actual chemical reaction of breaking that sugar apart that causes directional motion. Lysozymes are amazing. They are the free antibiotics you have in your tears. But, what about those computing proteins we talked about earlier? Computation has to do with information. Surely, information processing is far too sophisticated, far too complex of a process to be powered by normal Brownian motion, the random bumping into each other of particles, right? Well, check out the surface of DNA. Here's what a double-stranded chain of DNA looks like if we color its surface according to surface charge. It is red, which means it is negatively charged. Now, if we look at the lac repressor protein, it's the one we looked at earlier in this video, the computing protein we looked at earlier. Cells make all sorts of regulatory proteins, regulatory proteins that bind to DNA. This particular protein, the lac repressor, is one found in milk-eating bacteria. Its function is to turn milk-digesting genes off when milk is not in the environment. The milk digestion proteins that those genes code for, they're obviously not needed if there's no milk around to digest. Look at the charge distribution on the lac repressor protein. Here we see that not only is the foot of this protein positively charged, it is blue, meaning it would be attracted to DNA's negative charge. Not only is the foot of this protein positively charged, it is perfectly shaped to attach at least loosely to DNA anywhere, any chain of DNA, any sequence of DNA. You can see [clears throat] this just by looking at it. It's the reverse shape of a double-stranded chain of DNA. It has the reverse impression of DNA's twist and everything. This nearly perfect shape allows it to attach to DNA at pretty much any location, but it attaches loosely. Brownian motion will cause it to scoot along the chain of DNA, but when it hits its actual target, the very specific sequence that it is evolved to attach to, it sticks hard. It sticks to that DNA, and it stops anything else from sticking to that DNA. It prevents the cell from being able to read the genes that are near where the lac repressor is bound. Remember, just like protein, DNA is also a chain. It's a chain of molecules called nucleotides. There are four different types of nucleotides, often labeled A, T, C, and G. They can be arranged in any type of sequence. This particular protein, the lac repressor, is highly [snorts] attracted due to its charge and shape to a specific sequence, T G A G C. That sequence has a unique shape and a unique charge, and the foot of the lac repressor protein has the inverse charge and the inverse shape of that sequence. And that's it. That's how it turns genes off. It's just floating around in the cell, going all over the place. When it hits DNA, it's guided by that DNA. It's now searching the length of the DNA chain until it finds that specific sequence, and then it clamps down. Okay, so that's how the lac repressor protein turns milk-processing genes off. It just sticks to a chunk of DNA near those genes, gumming up the DNA so the cell can't use it. But, what about when milk is around and the cell needs those milk-digesting proteins? How do those genes get turned back on? Well, as mentioned, bacteria have evolved many different types of gene repressor proteins. The simplest repressors work by just loosely attaching to their target. In these cases, Brownian motion, or sometimes the activity of rowdy protein neighbors, will sometimes knock them off the DNA. When this happens, the cell starts using the genes that the repressor was blocking until the repressor manages to reattach. The lac repressor does work this way, too. It can simply fall off of its target
Segment 6 (25:00 - 30:00)
randomly, but it also has evolved a very special trick. When you look at the lac repressor protein, you find there are these little tiny grooves in it. Those are the nearly perfect size and shape for milk sugars to attach. If the cell is bathed in milk, those sugars are going to end up getting inside the cell, and there is a special version of those sugars, a partially processed that is extremely good at binding to the lac repressor. And when it binds to the lac repressor, so that lac repressor is currently attached to the DNA, when the sugar binds to the lac repressor, that changes the shape of the entire protein. It releases the DNA and drifts off again. This is an amazing system. The very type of food particle that those genes work to digest, that very same food particle is used to knock the repressor off of those genes, turning them on. They are only used, almost only used, when needed. And it's just because of the shape and the charge of the protein that attaches to those genes. Here we see that protein, just a normal physical molecule, through Brownian motion, it is Brownian motion that brings those little milk molecules into the lac repressor, causes it to release. The normal process of Brownian motion, combined with the physical shape and properties of protein, that's all you need to get computing to happen. Now, of course, it's not just any protein that will do that job. There is more than one way to skin a duck, and of course, there is more than one way to regulate a gene, but you do need a fairly specific protein shape. So, you need a very specific amino acid sequence that's able to do all of these things, and that is where evolution comes in. This is why I say that proteins are evolution's squishy magnetic Legos. A typical bacterial cell has roughly 4,000 protein-coding genes, and in most cases, each of those protein-coding genes codes for one specific type of protein. A gene is a stretch of DNA, a sequence of A's, C's, T's, and G's that code for a protein. They act as a template for a protein. Remember, a protein is a chain. DNA is also a chain, and it's because of that DNA can act as a template for a specific protein. It turns out that every three DNA nucleotides tell the cell which of the 20 different types of amino acids should be used next when it's building a protein. So, the sequence of a gene maps directly to the amino acid sequence in a protein. The gene, you could say, serves as the cell's memory. It can be used over and over and over again to keep on making that same protein. If a mutation occurs in DNA, that will change the amino acid sequence. Changing the amino acid sequence of a protein changes its folded shape, changes its function, and this is how you get evolution over time. A mutation in a lysozyme gene, for example, can modify its shape dramatically. A mutation in just the right place could break the gene, or a mutation can change its shape and charge slightly, just gently modifying how strong it binds to its target, or modifying what type of target it sticks to. All of the principles of evolution we've learned about so far in this series. So, optimization, co-option, duplication and recruitment. All of these principles work as perfectly on proteins and on protein complexes as they do on the anatomical structures we've studied so far. In fact, because proteins are so good at self-assembly, evolution through recruitment is far easier at the protein level than it typically is at the level of anatomy. So, in summary, a string of DNA acts as a template for a string of amino acids. acids, when it's built by the cell, it bends and it folds and it sticks to itself in a specific three-dimensional shape. A folded protein has charges on its surface. Some parts are negative, some parts are positive, some parts are neutral. A typical protein has parts that are highly stable. There's lots of bonds holding it together, and there are other parts that are highly mobile. They can bend, they can fold, they can twist. The normal process of Brownian motion allows individual proteins to self-assemble automatically into larger three-dimensional structures. Last but not least, the full process of evolution works all the way down at the level of proteins. And because proteins are so good at finding partners and self-assembling, no matter where the genes that coded for them exist in the cell's genome, evolution by recruitment, the merging together of previously separate parts. Evolution by recruitment is far easier at the level of protein than it is at the level of animal anatomy. In the next video, we're going
Segment 7 (30:00 - 31:00)
to look at the bacterial flagellum itself, how it assembles, how it is built inside the cell and outside the cell. We're going to look at how it functions, and then our final video will be how the bacterial flagellum evolved deep dive. I will be putting everything together, all the cool stuff we've learned about evolution, about co-option, about recruitment. Make sure you are subscribed to this channel. Make sure you click the bell icon so you do not miss one of these. If you are enjoying this series, please consider supporting me on patreon, patreon. com/statedclearly. Right now, over there, I just released a bonus video for patrons where you can see how I made this little protein complex, and you get to see the first time that I actually experimented with it and got it to self-assemble. It was super fun doing that. If you like this graphic that I kept showing during the video, there's a link to it down in the video description. It was made by David Goodsell, Alexander Rose, Maria Voit, and Rob Lowe. Huge thanks to them, and big, big thanks to Peter from the Clockwork Biology YouTube channel. He let me use his amazing protein animations. If you want to really understand how proteins work, I mean, next-level stuff, visit his channel. He also has a Patreon page, patreon. com/clockworkshow. I am very grateful to Chris Spronk and Bart Von Rossum. They are the ones that made this excellent simulation of protein folding.